
Przeciwciała, inaczej immunoglobuliny, to glikoproteiny o zdolnościach swoistego wiązania antygenów. Są najważniejszymi cząsteczkami układu odpornościowego. U człowieka wyróżnia się pięć klas przeciwciał, są to: IgG, IgA, IgM, IgD, IgE. Podział ten warunkują głównie różnice w budowie łańcuchów ciężkich poszczególnych klas.
Wszystkie przeciwciała zbudowane są z czterech łańcuchów polipeptydowych: dwóch lekkich (L) (4) i dwóch ciężkich (H) (3). W obu można wyróżnić, leżące na N-końcu, części zmienne V i na C-końcu części stałe C. Sekwencja aminokwasów w części zmiennej jest inna dla każdej cząsteczki przeciwciała. Łańcuchy lekkie połączone są z ciężkimi mostkami dwusiarczkowymi, natomiast na wysokości tak zwanego, regionu zawiasowego (6) łączą się ze sobą oba łańcuchy ciężkie, również za pomocą mostków dwusiarczkowych, ale także hydrofilowych i hydrofobowych wiązań niekowalencyjnych.
Oprócz immunoglobulin występujących w formie monomerycznej (posiadającej cztery łańcuchy), występują też formy polimeryczne, np. dimery ( IgA wydzielnicza) i pentametry (IgM). Formy te posiadają łańcuch łączący J, który może brać udział w scalaniu podjednostek.
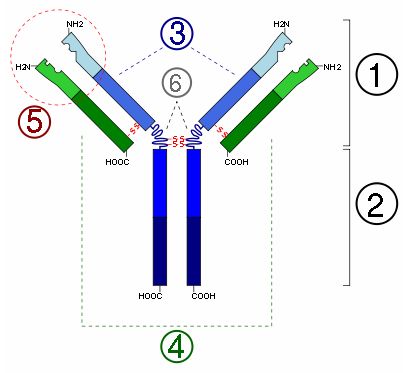
Rys.1 Budowa przeciwciał (opis w tekście)
W zależności od użytego enzymu (papaina, pepsyna) przeciwciało rozpada się na odpowiednie fragmenty. Trawienie papainą daje nam dwa takie same fragmenty Fab (1), zawierające miejsca wiążące antygen (5) oraz fragment aktywny biologicznie Fc (2), który jest odpowiedzialny za aktywację dopełniacza. W wyniku hydrolizy pepsyną otrzymujemy fragmenty F(ab’)2 i Fc’. Oprócz tego można wyróżnić fragment Fv, który zawiera wyłącznie części zmienne łańcuchów lekkiego i ciężkiego.
Mówiąc o immunoglobulinach trzeba wspomnieć o ich swoistości, powinowactwie, awidności i wartościowości.
Swoistość przeciwciała jest warunkowana przez przestrzenne ułożenie części zmiennych. Są one inne dla każdej klasy immunoglobulin wiążących różne antygeny. W danej grupie przeciwciał mogą wystąpić także różnice w sile wiązania determinanty antygenowej, czyli powinowactwie. Im bardziej dopasowane są miejsce wiążące antygen i epitop tym większa siła wiązania. W łączeniu tym występują: siły Van der Waalsa, siły elektrostatyczne, oddziaływania hydrofobowe oraz wiązania wodorowe. Wartościowość jest określana przez liczbę determinant antygenowych, które może związać pojedyncza cząsteczka przeciwciała. I tak, immunoglobuliny IgG, IgE oraz IgD posiadając dwa miejsca wiążące mogą związać dwie cząsteczki antygenu, a więc są dwuwartościowe, dimerczna forma IgA posiada cztery fragmenty Fab i jest czterowartościowa, pentametr IgM jest dziesięciowartościowy. Awidność zależy od powinowactwa i określa siłę wiązania antygenu posiadającego różne determinanty antygenowe przez przeciwciała.
Charakterystyka klas
IgA
Są pierwszą linią obrony błon surowiczych i śluzowych przed inwazją i kolonizacją mikroorganizmów. W układach tych występują w formie dimeru i zwierają tak zwany fragment wydzielniczy. W surowicy przeciwciała te są monomerami. Większość immunoglobulin IgA wytwarzana jest miejscowo i wydalana z wydzielinami, dlatego szacuje się, że organizm wytwarza ich więcej niż wszystkich innych klas. Istnieją dwie podklasy: IgA1 i IgA2.
IgE
Stężenie tych immunoglobulin jest w surowicy najmniejsze, ale odgrywają one znaczną rolę, głównie w reakcjach alergicznych organizmu. Po stymulacji antygenem łączą się z receptorami dla fragmentu Fc na komórkach tucznych i przy ponownym ataku tego antygenu powodują uwolnienie zawartości ziaren tych komórek. W swojej budowie nie posiadają regionu zawiasowego i odcinków ogonowych, co odróżnia je od IgM.
IgD
Występują głównie na powierzchni limfocytów i pełnią tam funkcję receptorów dla antygenu. Posiadają bardzo długi region zawiasowy, dzięki czemu wykazują dużą ruchliwość fragmentów Fab względem siebie.
IgG
Najwięcej immunoglobulin IgG występuje w surowicy, gdzie w stosunku do innych klas mają najwyższe stężenie. Są główną linią obrony przed mikroorganizmami w tkankach. Wyróżnia się cztery podklasy: IgG1, IgG2, IgG3, IgG4. Przeciwciała te (oprócz IgG4) inicjują klasyczną drogę aktywacji dopełniacza, co pozwala na zabicie rozpoznanych przez nie zakażonych komórek i mikroorganizmów. Wyróżniającą tę klasę immunoglobulin cechą jest to, że jako jedyna jest transportowana przez łożysko z organizmu matki do płodu. Zapewnia to odporność zarówno płodowi jak i niemowlęciu do około 3 miesiąca życia.
IgM
Są to pierwsze wytwarzane przeciwciała w rozwoju osobniczym i pierwsze uwalniane w trakcie odpowiedzi immunologicznej. Ich powinowactwo do antygenu jest raczej niskie, ale forma pentametru pozwala na wiązanie epitopu z dużą awidnością. Wolna cząsteczka IgM będąca pentametrem przypomina kształt płatka śniegu, natomiast wiążąc się wszystkimi podjednostkami z determinantami antygenowymi przybiera kształt tzw. kraba. Taka zmiana położenia uwidacznia fragmenty Fc, co sprzyja aktywowaniu się dopełniacza. Formy pojedynczych cząsteczek IgM występują na powierzchni limfocytów B i pełnią funkcję receptorów.
Kompleksy immunologiczne
Kompleksy immunologiczne to kompleksy antygen-przeciwciało. Każdy antygen zwykle zawiera kilka epitopów, a każde przeciwciało ma na ogół co najmniej dwa miejsca wiążące. Jeżeli obie cząsteczki wystąpią w odpowiednich ilościach, to połączenia między nimi mogą doprowadzić do powstania trójwymiarowej sieci, tak zwanego agregatu. Proces ten nazywa się precypitacją, czyli wtrącaniem się kompleksu z roztworu. W przypadku nadmiaru antygenu lub przeciwciała tworzą się rozpuszczalne agregaty. Kompleksy immunologiczne mogą stymulować odpowiedź odpornościową, ale ich odkładanie się w drobnych naczyniach może prowadzić też do stanów patologicznych np. kłębuszkowego zapalenia nerek.
Autor: Katarzyna Sala, studentka III roku biotechnologii na Katolickim Uniwersytecie Lubelskim
Literatura:
1. P.M. Lydyard, A. Whelan, M.W. Fanger ‘Immunology’
2. J. Gołąb, M. Jakóbisiak, W. Lasek ‘Immunologia’