
Autor: Anna Kurcek
Nośnikiem informacji genetycznej jest cząsteczka kwasu dezoksyrybonukleinowego – DNA. Tworzy ona w jądrze komórkowym delikatną, splątaną nitkę, czyli chromatynę, która podczas podziałów komórkowych ulega spiralizacji w struktury zwane chromosomami. Ich ilość i wielkość jest zawsze taka sama w obrębie gatunku, dzięki czemu spokrewnione osobniki są do siebie podobne. Wyjątkiem są tzw. mutanty, czyli organizmy, których genotyp uległ nagłemu i skokowemu przeobrażeniu – mutacji.
Zachodzące w sekwencji nukleotydowej zmiany mogą wywoływać różny wpływ na funkcjonowanie organizmu ułatwiając mu przystosowanie się do panujących w środowisku warunków (mutacje korzystne) lub zmniejszając jego szanse na przeżycie (warunkowo letalne), a nawet powodując jego śmierć (letalne). Należą one zatem do czynników warunkujących zmienność żywych organizmów oraz uważane są za jeden z motorów napędowych ewolucji. Zachodzące zmiany najczęściej nie wywierają żadnego wpływu na fenotyp osobnika. Są to tzw. mutacje ciche, nieme, zachodzące głównie w niekodujących obszarach DNA. Zmiany obszarów pozagenowych mogą jednak zaburzać pracę promotorów oraz sekwencji regulatorowych. Określając wpływ mutacji trzeba również pamiętać, że może się ona ujawnić z opóźnieniem oraz, że występujące zmiany mogą wpływać na siebie nawzajem.
Mutacje mutatorowe zwiększają częstość mutacji spontanicznych różnych genów, natomiast mutacje antymutatorowe, obniżają częstość występowania zmian dzięki poprawie zdolności korektorskich polimerazy. Wyróżnia się także tzw. gorące miejsca (ang. hot spots), w których zaburzenia występują z większą częstotliwością. Hipermutacją nazywamy zaś zjawisko zwiększenia częstości mutacji w komórce zwierzęcej.
Powstałe zmiany mogą prowadzić do odtworzenia formy wyjściowej allelu. Zjawisko takie nazywa się rewersją. Odwrócenie skutków mutacji może być również wynikiem tzw. mutacji supresorowych, zachodzących w innym miejscu niż zmiana pierwotna.
Najistotniejsze są mutacje zachodzące w komórkach rozrodczych – germinalne. Tylko one przekazywane są następnym pokoleniom. Zmiany w komórkach somatycznych mogą prowadzić do transformacji nowotworowej lub wpływać na rolę białek powodując utratę ich aktywności lub nabycie nietypowych funkcji. Dotyczą jednak jedynie osobnika u którego wystąpiły. Ze względu na podatność na mutacje, geny dzielimy na labilne (gen VIII czynnika krzepliwości krwi), w których zmiany powstają de novo oraz stabilne (gen pląsawicy Huntingtona), których mutacja jest głównie przekazywana z pokolenia na pokolenie.
Rodzaje mutacji
Podziału mutacji dokonuje się najczęściej na podstawie wielkości zmian zachodzących w DNA.
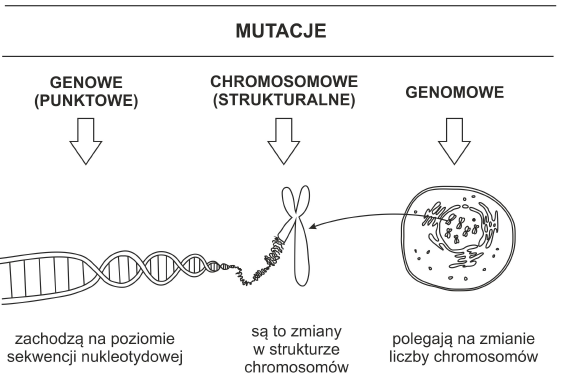
Rys. 1. Rodzaje mutacji.
Mutacje punktowe określane też jako mutacje genowe dotyczą sekwencji nukleotydowej. Mogą one polegać na zastąpieniu jednej zasady azotowej inną. Tego typu podstawienie nazywa się substytucją lub bardziej dokładnie tranzycją (w przypadku, gdy puryna zastępowana jest inną puryną lub pirymidyna pirymidyną), bądź transwersją (zamiana zasady purynowej na pirymidynową lub odwrotnie). Prowadzą one do zmian w sekwencji aminokwasowej kodowanego białka. Mogą zatem przyczyniać się do upośledzenia jego funkcjonowania oraz zmiany budowy poprzez skrócenie łańcucha (mutacja nonsens) lub jego wydłużenie (zmiany kodonu terminacyjnego).
Wyjątkiem jest tzw. mutacja synonimiczna, w efekcie której powstaje trójka nukleotydów kodująca ten sam aminokwas, co pierwotny kodon. Jest to możliwe dzięki zdegenerowaniu kodu genetycznego.
Mutacje punktowe obejmują również utratę (delecję) oraz wstawienie (insercję) jednego lub kilku nukleotydów. Tego typu zaburzenia powstają głównie w wyniku tzw. poślizgu replikacyjnego fragmentów zawierających krótkie, powtórzone sekwencje i prowadzą do zmiany fazy odczytu kodowanych aminokwasów, czego efektem jest powstawanie zupełnie innego łańcucha polipeptydowego.
Aberracje chromosomowe – związane ze strukturą chromosomów
Występują najczęściej podczas podziałów komórkowych jako efekt pęknięcia chromosomu, nieprawidłowego podziału centromeru lub błędów zachodzących podczas wymiany crossing-over. Do zmian tych zaliczamy:
• Delecję, czyli wypadnięcie fragmentu chromosomu z części dystalnej (deficjencja) lub środkowej (delecja interstycjalna). Utrata zbyt dużej ilości materiału genetycznego (powyżej 3%) powoduje śmierć zmutowanego osobnika.
• Duplikację, która polega na zwielokrotnieniu fragmentu chromosomu. Zazwyczaj nie zaburza ona funkcjonowania organizmu lecz prowadzi do utworzenia tzw. pseudogenów.
• Inwersję, powstającą na skutek nieprawidłowej naprawy pękniętych fragmentów chromosomu, które ulegają odwróceniu o 180°. Zmiana pozycji genów może wpływać na ich ekspresję. Wyróżnia się dwa typy tej mutacji: inwersję pericentryczną, obejmującą fragment chromosomu zawierający centromer, paracentryczną – dotyczącą odcinka bez centromeru.
• Translokacja, polega na przeniesieniu fragmentu jednego chromosomu na drugi chromosom – niehomologiczny (interchromosomalna, zewnętrzna) lub homologiczny (intrachromosomalna, wewnętrzna). Wyróżnia się też translokację wzajemną, podczas której dochodzi do wzajemnej wymiany odcinków oraz transformację robertsonowską (fuzję centryczną), prowadzącą do utraty materiału genetycznego znajdującego się w krótkich ramionach dwóch łączących się ze sobą chromosomów.
• Podział centryczny, czyli nieprawidłowy podział centromeru zachodzący w chromosomie o dwóch ramionach. Prowadzi to do powstania pary chromosomów teocentrycznych oraz zwiększenia ich liczby w komórce.
• Chromosom kolisty tworzy się podczas utraty końcowych odcinków chromosomu i połączeniu się ze sobą powstałych w ten sposób zakończeń.
• Izochromosom, czyli chromosom składający się z samych długich lub jedynie krótkich ramion. Jego obecność powodowana jest poprzecznym podziałem centromeru.
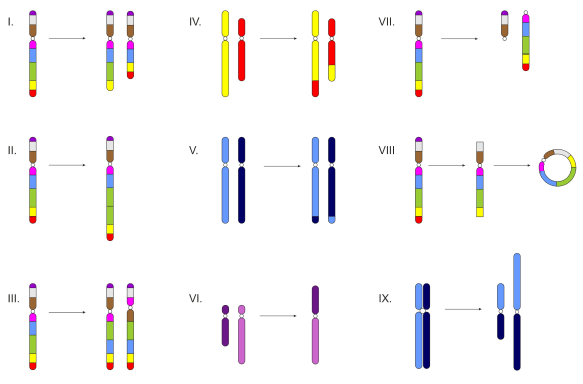
Rys. 2. Mutacje chromosomowe.
I. delecja 1. terminalna, 2. interstycjalna; II. duplikacja; III. inwersja 1. paracentryczna, 2. pericentryczna; IV. translokacja interchromosomalna (zewnętrzna); V. translokacja intrachromosomalna (wewnętrzna); VI. fuzja centryczna (transformacja robertsonowska); VII. podział centryczny; VIII. chromosom kolisty; IX. izochromosom.
Mutacje genomowe które polegają na zmianie liczby chromosomów
Odchylenia dotyczące poszczególnych par chromosomów homologicznych nazywa się aneuploidiami. Powstają one na skutek uszkodzenia wrzeciona kariokinetycznego podczas podziałów komórkowych i prowadzą do zwiększenia lub zmniejszenia liczby chromosomów. Wielkość zmian warunkuje typ mutacji, spośród których wyróżnia się:
nullisomie (2n-2),
monosomie (2n-1),
trisomie (2n+1),
tetrasomie (2n+2),
podwójne monosomie (2n-1-1),
podwójne trisomie (2n+1+1).
Zaburzenia mogą również dotyczyć całego garnituru chromosomów i prowadzić do zmiany ploidalności organizmów. Takie mutacje nazywa się euploidiami. Mogą one tworzyć się wskutek nieprawidłowych podziałów komórkowych i objawiać zwielokrotnieniem homologicznego zestawu genów u jednego organizmu – autopoliploidu. Allopoliploidem nazywamy zaś organizm u którego zwielokrotniony garnitur chromosomowy ma pochodzenie mieszańcowe. Podwojenie niehomologicznych zestawów chromosomów powstaje w wyniku połączenia się gamet pochodzących od osobników należących do blisko spokrewnionych ze sobą gatunków.
Przyczyny powstawania mutacji w DNA
Mutacje mogą zachodzić spontanicznie, jako skutek pomyłek zachodzących podczas replikacji. Przyczyną takich zmian bywają błędy polimerazy, która ze względu na obniżoną skuteczność selekcji nukleotydów oraz ograniczone zdolności naprawcze wbudowuje w nowozsyntetyzowaną nić niekomplementarne cząstki. Innym powodem jest tzw. tautomeria zasad azotowych, czyli występowanie form iminowych (z grupą iminową =NH zamiast aminowej –NH2) oraz enolowych (z grupą enolową =C-OH zamiast ketonowej –CO), które choć nie zaburzają struktury podwójnej helisy, tworzą wiązania z innymi niż normalnie zasadami (tymina łączy się w parę z guaniną, adenina zaś z cytozyną) i przyczyniają się do powstawania drobnych mutacji punktowych.
Zmiany w informacji genetycznej mogą być również indukowane, tzn. wywołane obecnością czynników mutagennych, działających na cząsteczkę DNA w sposób pośredni lub bezpośredni. Siłę mutagenu, czyli jego genotoksyczność określa się za pomocą Testu Amesa. Wykorzystuje on szczepy komórek bakteryjnych Salmonella typhimurium z mutacją blokującą produkcję histydyny. Częste mutacje prowadzą do rewersji i przywrócenia zdolności tworzenia tego niezbędnego aminokwasu. Ilość wyhodowanych bakterii pozwala zatem oznaczyć natężenie efektu mutagennego badanego czynnika.
Czynniki fizyczne jako mutageny
• Promieniowanie ultrafioletowe (UV) – słabo przenika przez tkanki, jednak jest bardzo silnie pochłaniane przez DNA. Prowadzi do dimeryzacji zasad azotowych. Zmiana ta obejmuje głównie sąsiadujące ze sobą pirymidyny (najczęściej tyminy), które łączą się ze sobą silnymi wiązaniami kowalencyjnymi, zaburzając tym samym proces replikacji oraz prowadząc do powstawania delecji. Tego typu promieniowanie może przyczyniać się również do rozrywania podwójnej helisy DNA oraz hydratacji cytozyny i uracylu.
• Promieniowanie jonizujące: (X, α, β, γ, kosmiczne) – bardzo dobrze przenika przez tkanki. Powoduje wybicie elektronów z atomów i cząsteczek, a powstałe w ten sposób jony mogą inicjować różne reakcje chemiczne zachodzące w komórkach prowadzić do produkcji mutagennych nadtlenków organicznych oraz wolnych rodników, których rozpad wyzwala znaczne ilości energii wywołujące zmiany chemiczne w DNA. Rodzaj i natężenie promieniowania, warunkuje wielkość zmian, które mogą obejmować mutacje punktowe, delecje, insercje oraz pękanie łańcuchów DNA wywołane rozerwaniem wiązań wodorowych. Może to być przyczyną powstawania transformacji nowotworowych oraz mutacji uniemożliwiających replikację genomu. Im większa zawartość tlenu cząsteczkowego w środowisku, tym silniejszy jest efekt mutagenny promieniowania jonizującego. Długotrwałe stosowanie małych dawek promieniowania stwarza większe możliwości naprawy DNA, jest zatem mniej niebezpieczne niż jednorazowe, duże napromieniowanie. Trzeba jednak pamiętać, że procesy reparacyjne zachodzą tylko w komórkach aktywnych metabolicznie, dlatego komórki nieaktywne, np. plemniki są bardziej zagrożone mutacją. Sprawność mechanizmów naprawczych jest różna u różnych gatunków organizmów. Komórki ulegające szybkim podziałom (np. embrionalne) są bardziej wrażliwe na skutki promieniowania, niż komórki dzielące się wolno (np. neurony, komórki mięśniowe) oraz nie dzielące się wcale.
• Protony i neutrony uwalniane przez radioaktywne izotopy pierwiastków (np. kobalt-90, 32P).
• Temperatura – jej nagłe zmiany mogą prowadzić do hydrolizy wiązania β-N-glikozydowego, łączącego zasadę azotową z cukrem. Mutacja ta najczęściej dotyczy puryn. Pozbawiony zasady nukleotyd jest niestabilny i ulega szybkiej degradacji tworząc tzw. miejsce AP (apurynowe, apirymidynowe). Powstała w ten sposób luka zwykle jest uzupełniana. Jednak, gdy w komórce włączony jest system naprawy SOS, wszystkie przerwy w podwójnej helisie wypełniane są adeniną, co prowadzi do powstawania mutacji.
Czynniki chemiczne jako mutageny
• Czynniki deaminujące – usuwają grupę aminową z zasad azotowych; Przykładem może być kwas azotawy (HNO2), który przekształca guaninę w ksantynę, adeninę w hipoksantynę, a cytozynę w uracyl. Efektem tych zmian jest zablokowanie replikacji lub tranzycja nukleotydów.
• Analogi zasad (5-bromouracyl, 2-aminopuryna), ze względu na duże podobieństwo do występujących w nukleotydach zasad azotowych mogą być one wbudowywane do DNA podczas replikacji. Ich obecność zmniejsza stabilność wiązań wodorowych oraz prowadzi do nieprawidłowego parowania zasad.
• Czynniki alkilujące (iperyty, tlenki etylenu, halogenki metylu, etylonitrozomocznik, produkty metabolizmu azotynów), czyli związki, które dodają do nukleotydów grupy alkilowe lub arylowe. Efekt ich działania jest różny i zależy od pozycji w której zostaje zmodyfikowany nukleotyd oraz od rodzaju dodawanej grupy. Metylacja prowadzi do mutacji punktowych, gdyż osłabia zdolność do tworzenia wiązań komplementarnych. Dodanie innych grup hamuje replikację DNA.
• Hydroksylamina (NH2OH) wchodzi w reakcję z cytozyną i zamienia ją na związek podobny do uracylu, co z kolei prowadzi do poreplikacyjnej tranzycji w tyminę. Działa również na enzymy komórkowe.
• Czynniki interkalujące (barwniki akrydynowe, np. bromek etydyny) – wnikają w łańcuch DNA i powodują rozsunięcie się zasad azotowych, deformując tym samym strukturę podwójnej helisy. Prowadzi to do błędów w replikacji, delecji, insercji oraz zmiany ramki odczytu.
• Reaktywne formy tlenu, czyli takie, które zawierają niesparowany elektron. Niektóre z nich są naturalnymi produktami metabolizmu, a ich podwyższona zawartość nazywana jest stresem oksydacyjnym.
• Policykliczne węglowodory aromatyczne (benzopiren), występujące w dymie papierosowym, spalinach samochodowych. Prowadzą do powstawania tranzycji tpu GC -> TA.
• Niektóre leki psychotropowe, cytostatyki, antybiotyki.
• Konserwanty, środki ochrony czystości, środki chwastobójcze, owadobójcze.
Czynniki biologiczne mogące wywoływać mutacje
• Niektóre wirusy (różyczki, opryszczki).
• Pierwotniaki wywołujące toksoplazmozę.
• Mykotoksyny produkowane przez niektóre grzyby pleśniowe (gł. aflatoksyny).
Literatura:
„Genetyka ogólna” Skrypt do ćwiczeń dla studentów biologii; Anna Sadakierska-Chudy, Grażyna Dąbrowska, Anna Goc; Uniwersytet Mikołaja Kopernika Toruń 2004;
„Genetyka zwierząt” Przewodnik do ćwiczeń; Wilhelm Grzesiak, Arkadiusz Marian Kawęcki; Akademia Rolnicza w Szczecinie, Szczecin 1998;
„Biologia vademecum maturzysty”; Ewa Pyłka-Gutowska, Wydanie siódme zmienione, Wydawnictwo „OŚWIATA”, Warszawa 2002;