
Fotosynteza to synteza związków organicznych (glukozy) z prostych związków nieorganicznych (sole mineralne, woda) w obecności odpowiednich barwników i przy udziale energii świetlnej. W procesie tym powstają związki mniej utlenione, a co za tym idzie mające wyższą wartość energetyczną. Stanowią one źródło budulca dla autotrofów oraz są pokarmem dla heterotrofów. Dzięki wiązaniu CO2 i uwalnianiu tlenu, fotosynteza zapewnia również utrzymanie równowagi gazowej atmosfery. Jest to zatem podstawowy proces biologiczny warunkujący życie na Ziemi.
Fotosynteza zachodzi u roślin, bakterii i pierwotniaków i zlokalizowana jest w specjalnych obszarach komórki. U roślin są nimi chloroplasty, czyli organelle skupione w tkance miękiszowej liści. Fotosyntetyzujące Procaryota posiadają ciałkach chromatoforowe (barwnikowe) o budowie lamelarnej, natomiast u sinic znajdujemy tylakoidy oddzielone od błony cytoplazmatycznej.
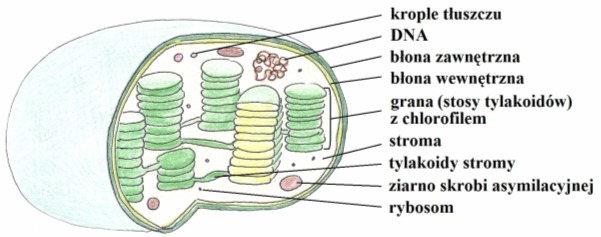
Rys. 1. Budowa chloroplastu
Fotosynteza jest złożonym procesem, który zachodzi w dwóch fazach.
Faza jasna (świetlna)
To szereg reakcji fotochemicznych – wymagających światła do swojego przebiegu. Zachodzi w granach, gdzie zgromadzony jest chlorofil. Istotą tej fazy jest przekształcenie energii świetlnej w energię wiązań chemicznych, produktami jest siła asymilacyjna w postaci NADPH2 i ATP oraz tlen jako produkt uboczny.
Biorą w niej udział dwa fotosystemy chlorofilu: ewolucyjnie starszy PSI (chlorofil a i karotenoidy; 700nm) i PSII (chlorofil a i b, ksantofil; 680nm). Różnią się one między sobą głównie właściwościami występujących w nich cząsteczek chlorofilu i karotenoidów. Oba usytuowane są w innych częściach gran i działają razem, choć inaktywacja jednego nie wstrzymuje fotosyntezy.
Pochłaniane przez chlorofil fotony powodują wybicie elektronów z cząsteczki chlorofilu. Są one natychmiast wychwytywane przez przenośniki elektronów takie jak NADP, ferredoksynę, czy cytochromy. Transportowany elektron stopniowo traci energię, która częściowo rozpuszcza się w postaci ciepła, a częściowo magazynowana jest w postaci ATP. Proces ten nazywa się fosforylacją fotosyntetyczną i może być:
• cykliczny – gdy elektrony PSI po przejściu przez szereg przenośników wracają do fotosystemu I;
• niecykliczny – gdy elektrony wybite z PSI transportowane są na przenośnik wodoru NADP+, a powstałą w chlorofilu lukę wypełniają elektrony z innej cząsteczki chlorofilu (PSII). Fotosynteza niecykliczna związana jest z fotolizą wody, czyli jej rozkładem pod wpływem światła:
H2O-> 2H+ + 4e– +O2
Kationy wodoru łącza się z NADP tworząc NADPH2, który wraz z ATP stanowi siłę asymilacyjną wykorzystywaną następnie w fazie ciemnej. Elektrony pochodzące z cząsteczki wody wędrują na chlorofil wypełniając powstałe w nim luki (PSII przekazuje je PSI). Natomiast tlen poprzez aparaty szparkowe wydostaje się do atmosfery.
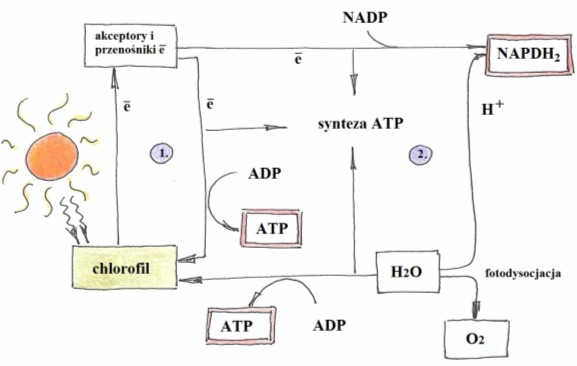
Rys. 2. Faza jasna
Faza ciemna (cykl Calvina)
Jest to cykl redukcji CO2. Składa się z cyklu reakcji biochemicznych wykorzystujących energię powstałą kosztem światła. Zachodzi w stromie chloroplastów, gdzie zgromadzone są wszystkie potrzebne do jej przebiegu enzymy.
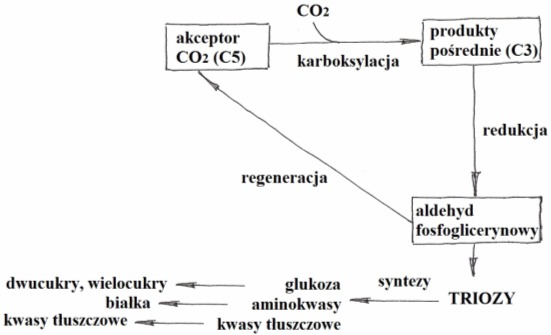
Rys. 3. Faza ciemna
Wyróżnia się trzy główne etapy fazy ciemnej:
• karboksylację: pobrany z atmosfery CO2 przyłączany jest do uaktywnionego pięciowęglowego cukru – rybulozodifosforanu (RuDP) lub rybulozy (RDP). Po przyłączeniu związek ten, już jako sześciowęglowy rozpada się na dwie cząsteczki o 3 atomach węgla – kwasy fosfoglicerynowe (PGA);
• redukcję: dotyczy redukcji PGA do 6 cząsteczek aldehydu 3-fosfoglicerynowego (GAP). Reakcja odbywa się z udziałem siły asymilacyjnej w postaci NADPH2 i ATP;
• regenerację: podczas której pięć cząsteczek GAP wykorzystywanych jest do odbudowy (regeneracji) akceptora CO2 – RuDP (jednocześnie przyłączane są 3 cząsteczki CO2). Pozostała jedna cząsteczka stanowi produkt netto fotosyntezy i jest substratem służącym do wytwarzania bardziej złożonych związków organicznych.
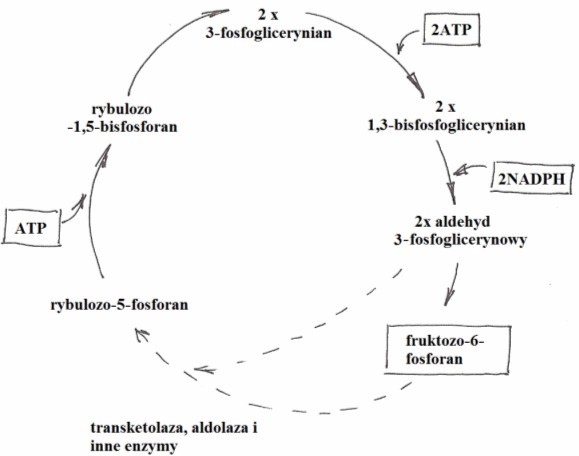
Rys. 4. Cykl Calvina
Większość roślin asymiluje CO2 z atmosfery w sposób bezpośredni przez włączenie go w redukcyjny cykl Calvina. Akceptorem CO2 jest pięciowęglowy RuDP, a pierwszym trwałym produktem trójwęglowy kwas 3-fosfoglicerynowy (PGA). Jest to tak zwana fotosynteza typu C3.
U niektórych roślin (trzcina cukrowa, kukurydza) zachodzi fotosynteza typu C4.
Akceptorem CO2 jest związek trójwęglowy – kwas fosfoendopirogronowy (PEP), a produktem reakcji czterowęglowy kwas szczawiooctowy.
U tych roślin przyswajanie dwutlenku węgla zachodzi w dwóch etapach:
• wiązanie CO2 przez PEP, który w wyniku przemian przekształca się w kwas jabłkowy (komórki mezofilu liściowego, małe chloroplasty);
• w komórkach chlorenchymy następuje dekarboksylacja kwasu jabłkowego do kwasu pirogronowego i CO2, który zostaje włączony w cykl Calvina i wbudowywany w związki organiczne. Kwas pirogronowy wraca do mezofilu.
Wiązanie CO2 w pierwszym cyklu przemian umożliwia podwyższenie jego stężenia i przyspiesza fotosyntezę, znacznie podwyższając produktywność rośliny.
Intensywność fotosyntezy zależy od wielu czynników:
zewnętrznych:
• światło – limituje przebieg fazy jasnej fotosyntezy. Jest również głównym czynnikiem powodującym rozwój miękiszu zieleniowego liści, syntezę chlorofilu tworzenie się chloroplastów z proplastydów oraz ich rozmieszczenie w komórkach. Fotosynteza zachodzi zarówno przy oświetleniu naturalnym jak i elektrycznym. Jednak jedynie część energii padającej na liść ulega przetworzeniu w energię chemiczną asymilantów. Szybkość procesu zależy zarówno od barwy jak i intensywności światła oraz od typu rośliny. Cieniolubne, np. zasiedlające runo leśne, największą wydajność osiągają już przy 1/10 intensywności pełnego światła słonecznego. Rośliny światłolubne, np. zboża, rosną najlepiej przy intensywnym oświetleniu. Zbyt silne światło hamuje fotosyntezę powodując utlenienie i inaktywację cząsteczek chlorofilu. Wzrasta też nadmiernie transpiracja, komórki tracą turgor, przez co szparki zamykają się hamując dopływ CO2.
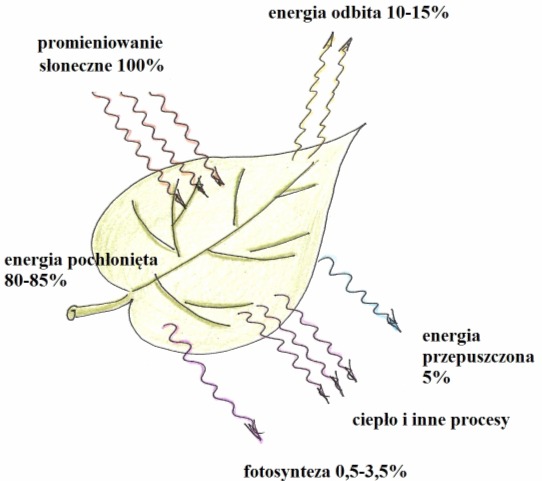
Rys. 5. Wykorzystanie energii świetlnej
• dwutlenek węgla (CO2) – występujące w atmosferze stężenie nie jest optymalne dla roślin i wynosi 0,034%. Jego wzrost do wartości 0,15% (efekt cieplarniany) podnosi trzykrotnie intensywność fotosyntezy. Z tego powodu w uprawie stosuje się nawozy organiczne – podczas ich rozkładu bakterie produkują dwutlenek węgla. Gazowy CO2 przedostaje się przez szparki do systemu przestworów międzykomórkowych, skąd dyfunduje do komórek miękiszu zieleniowego. Rośliny wodne pobierają go postaci jonów HCO3–.
• woda – substrat fotosyntezy. Jest czynnikiem decydującym o stopniu rozwarcia aparatów szparkowych. Uwadniając komórkę warunkuje procesy wymiany gazowej
• sole mineralne – źródło substancji do syntez, aktywatory wielu przemian. Niedostatek tylko jednego czynnika odżywczego, nawet przy wystarczającym dopływie pozostałych, ogranicza fotosyntezę (prawo minimum). W przypadku niedoboru soli azotu wstrzymana zastaje synteza chlorofilu i powstaje chloroza liści.
• temperatura – fotosynteza jest procesem enzymatycznym, którego optimum termiczne wynosi 20-30ºC. Po jej przekroczeniu obserwuje się szybki spadek intensywności zachodzących reakcji. Zakres tolerancji roślin waha się w dużym przedziale i zależy od ich gatunku i miejsca występowania. W większości przypadków przy temperaturze 40 ºC fotosynteza ustaje.
wewnętrznych:
• budowa organów i tkanek biorących udział w fotosyntezie: liście, układ chlorenchymy, aparaty szparkowe (ilość i rozmieszczenie), tkanka magazynująca wodę, barwniki;
• fizjologiczne właściwości rośliny:
o zdolność do manipulacji ułożeniem liści i chloroplastów;
o sprawność systemu dostarczającego wodę i sole mineralne;
o mechanizmy chroniące przed przegrzaniem i nadmierną transpiracją.
Rys. 6. Wpływ światła na rozmieszczenie chloroplastów w komórce
Fotoautotrofy wykorzystują energię promieniowania słonecznego o długości fali ok. 400 -700 nm, przy czym absorbują głównie fale o długości odpowiadającej barwie niebieskiej i czerwonej. Kluczową rolę podczas przebiegu tego procesu odgrywają barwne związki chemiczne zwane barwnikami fotosyntetycznymi (asymilacyjnymi). Występują one w większej ilości u roślin cieniolubnych niż u światłolubnych.
Wyróżnia się trzy główne grupy barwników:
Chlorofile – pochłaniają promienie o długości fali niebieskiej i czerwonej. Zielone barwniki rozpuszczają się w rozpuszczalnikach organicznych i tłuszczach, są jednak nierozpuszczalne w wodzie:
o chlorofil a – podstawowy, niebieskozielony barwnik, występuje u wszystkich fotoautotrofów z wyjątkiem bakterii;
o chlorofil b – zielonożółty barwnik występuje u roślin wyższych i glonów, niezależnie od chlorofilu a i w mniejszej ilości;
o chlorofil c występuje w niewielkich ilościach u okrzemek, brunatnic i niektórych wiciowców;
o chlorofilu d towarzyszy chlorofilowi a u krasnorostów;
o bakteriochlorofile.
Cząsteczka każdego chlorofilu zbudowana jest z feoporofiryny, czyli pochodnej porfiryny. Centralne miejsce w zajmuje atom magnezu łączący się z atomami azotu każdego z pierścieni. Barwniki zielone pochłaniają światło widzialne w zakresie 370-760 nm, przy czym każdy z nich ma swoje charakterystyczne widmo absorpcyjne z dwoma szczytami – jednym w zakresie czerwieni, drugim z części niebiesko – fioletowej.
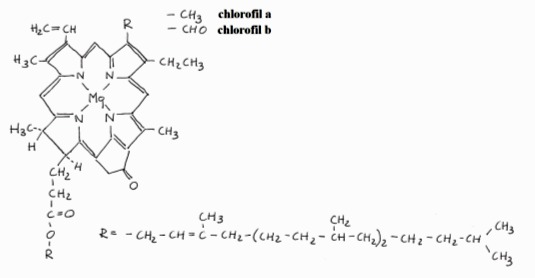
Rys. 7. Wzór strukturalny cząsteczki chlorofilu
Karotenoidy (np. β-karoten) – to żółte lub pomarańczowe barwniki pomocnicze, występujące u wszystkich fotoautotrofów, Absorbują światło z niebiesko – fioletowej części widma, a następnie przekazują ją na cząsteczkę chlorofilu. Ponadto chronią one fotosystemy przed nadmiarem docierającej energii świetlnej, którą pochłaniają lub przekierowują na inne procesy fizjologiczne. Zabezpieczają również komórkę przed reaktywnymi formami tlenu (aktywność przeciwutleniająca). Karotenoidy należą do tetraterpenów (40-węglowych terpenoidów). Ich cechą jest fotolabilność, czyli uleganie przemianom w obecności światła. Występują w stężeniu mniejszym niż chlorofil.
Fikobiliny (np. fikocyjan, fikoerytryna) – barwniki pomocnicze występujące u krasnorostów i sinic. Jako jedyne spośród barwników fotosyntetycznych powiązane są z rozpuszczalnymi w wodzie białkami. Wychwytują energię świetlną w obszarze 450-600 nm, a następnie przekazują ją na chlorofil. Jest to forma przystosowania się do życia na dużych głębokościach.
Autor: Anna Kurcek
Literatura:
• Lubert Stryer „Biochemia”, przekład zbiorowy pod redakcją Jacka Augustyniaka i Jana Michejdy z czwartego wydania amerykańskiego; Wydawnictwo Naukowe PWN, Warszawa 2003;
• Ewa Pyłka-Gutowska „Biologia. Vademecum Maturzysty”; Wydawnictwo „Oświata”; Warszawa 2002;
• Henryk Wiśniewski „Biologia dla klas III liceum ogólnokształcącego o profilu podstawowym i biologiczno-chemicznym”, AGMEN; Warszawa 1997.