
Chromosomy (z gr. chroma = kolor, soma = ciało) są jedną z form organizacji materiału genetycznego zgromadzonego w komórce. Tworzą się one podczas procesu, określanego jako kariokineza, na drodze spiralizacji włókien chromatyny jądra interfazowego. Najlepiej widać je w trakcie metafazy, podczas której osiągają maksimum kondensacji (u Eucaryota DNA upakowany jest wtedy około 10 tys. razy). Tak zagęszczony materiał genetyczny sprawia, że jest on nieaktywny transkrypcyjnie i replikacyjnie.
Struktura chromosomu
Podstawową jednostką strukturalną chromosomu jest włókno nukleosomowe – zbudowane z nuklosomów, czyli dwuniciowego DNA nawiniętego na białkowy rdzeń, który składa się z ośmiu białek histonowych (2x H2A, 2x H2B, 2x H3, 2x H4) i umożliwia wygięcie cząsteczki kwasu dezoksyrybonukleinowego oraz neutralizuje jej ujemny ładunek. Na jeden taki oktamer przypada 146 par zasad DNA tworzącego 1,8 zwoju. Znajdujący się poza rdzeniem histon H1 – stabilizuje strukturę włókna.
Nukleosomy powiązane ze sobą odcinkami DNA łącznikowego o długości od 0 do 80 par zasad. W ten sposób powstaje struktura przypominająca paciorki nawinięte na sznurek (ang. beads-on-a-string). Zwija się ona wokół niewidocznej osi tworząc w ten sposób strukturę zwaną solenoidem lub włóknem o średnicy 30 nm, która układa się w pętle budujące chromatydy. W miejscach zwanych MAR (ang. Matrix Associated Regions) pozwijane domeny łączą są z białkowym szkieletem chromosomu. Ten poziom organizacji DNA stabilizowany jest przez białka niehistonowe – głównie topoizomerazę II.
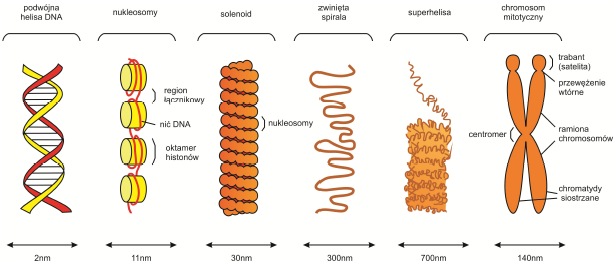
Rys. 1. Kolejne stopnie upakowania materiału genetycznego
Budowa chromosomu
Chromosomy zbudowane są z dwóch chromatyd siostrzanych połączonych ze sobą centromerem. Zlokalizowany jest on w tzw. przewężeniu pierwotnym, charakteryzującym się mniejszą niż reszta chromosomu średnicą.
DNA centromerowy stanowi zaledwie 1% całości jądrowego materiału genetycznego. Budująca go heterochromatyna składa się z tandemowo ułożonych, nietranskrybowanych sekwencji i nazywana jest satelitarnym DNA (satDNA).
W pobliżu centromerów znajdują się kinetochory, czyli białkowe struktury do których przyłączają się włókna wrzeciona kariokinetycznego. Przyjmują one głównie postać dyskowatej struktury ulokowanej na powierzchni każdej z chromatyd. U niektórych prymitywnych organizmów aktywność kinetochorowa może być rozproszona na całej długości chromosomów (chromosomy holokinetyczne , homocentryczne).
Chromosomy z jednym centromerem nazywa się monokinetycznymi lub inaczej monocentrycznymi. Ich większa liczba występuje rzadko i zakłóca podziały jądra komórkowego. Istnieją również obszary o wtórnej aktywności centromerowej, tzw. neocentromery, które pozbawione są przewężenia pierwotnego.
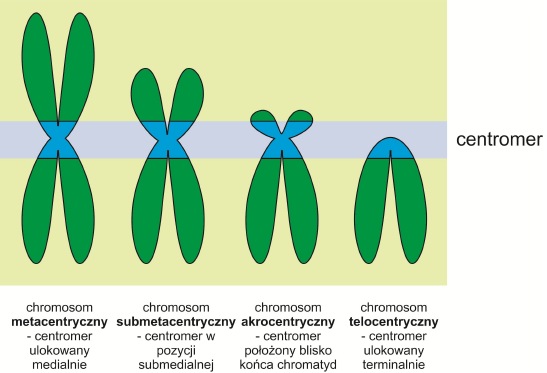
Rys. 2. Typy morfologiczne chromosomów
Strukturę chromosomów stabilizują telomery. Są one ulokowane na końcach chromatyd i skracają się podczas każdego podziału komórki. DNA telomerowy nie koduje żadnych genów i nie jest zorganizowany w struktury nukleosomowe. Zamiast białek histonowych występują tu specyficzne białka telomerowe tworzące wraz z DNA struktury zwane telosomami. Wykazują one ekstremalny konserwatyzm ewolucyjny.
Każda komórka posiada chociaż jedną parę chromosomów jąderkotwórczych. Zawierają one wyspecjalizowany obszar zwany NOR (ang. Nucleolar Organizer Region) – odcinek zawierający tandemowo ułożone geny rRNA (18S- 5,8S- 25S) warunkujące transkrypcję 45S prerybosomowego RNA. Podczas anafazy obszary te uczestniczą w tworzeniu jąderka – stąd pochodzi ich nazwa. Geny rRNA są jedynymi genami, które można zlokalizować w genomie na podstawie struktury chromosomu – dzięki obecności przewężenia wtórnego oddzielającego tzw. trabant, czyli satelitę.
Cytogenetyka
Kompletny zestaw chromosomów komórki somatycznej organizmu to kariotyp. Jest on charakterystyczny dla osobników tego samego gatunku i tej samej płci, a składają się na niego autosomy oraz chromosomy płci, czyli allosomy. U różnych organizmów liczba autosomów jest inna. U diploidów występują one w homologicznych parach, zaś u poliploidów jest ich odpowiednio więcej.
Kariotyp opisywany jest graficznie, w postaci idiogramów, stanowiących wynik badania cytogenetycznego. Przedstawiają one homologiczne pary chromosomów ułożone w kolejności od największego do najmniejszego i z centromerami na jednym poziomie. Chromosomy płci oraz NOR umieszczane są na końcu.
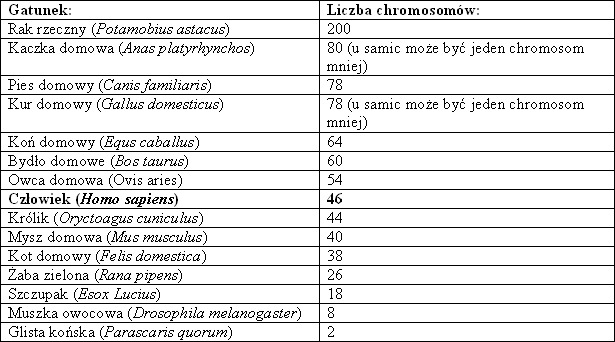
Tab. 1. Zestawienie diploidalnej liczby chromosomów w komórkach niektórych zwierząt i człowieka
Chromosomy różnią się od siebie długością, wielkością ramion, położeniem centromerów, stosunkiem długości ramienia długiego do krótkiego (indeksem ramion), stosunkiem długości ramienia krótkiego do całkowitej długości chromosomu (indeks centromerowy) oraz występowaniem przewężenia wtórnego. Cechy te nie zawsze umożliwiają jednak ich identyfikację. Z tego powodu stosuje się specjalne barwienia różnicowe, pozwalające uwidocznić wzór prążkowy chromosomu, tzn. specyficzny zestaw poprzecznych prążków różniących się od siebie grubością i intensywnością. Ich rozmieszczenie jest stałą i powtarzalną cechą wykorzystywaną również przy wykrywaniu mutacji.
Wyróżnia się cztery typy prążków:
– związane z organizatorem jąderka (NOR);
– związane z centromerem;
– prążki heterochromatynowe;
– prążki euchromatynowe.
Inna klasyfikacja prążków opiera się o metodę ich barwienia:
• barwione absorpcyjnie – z wykorzystaniem barwnika Giemsy:
– prążki C – ujawniane metodą BSG (Barium hydroxine/ Saline/Giemsa); reprezentują heterochromatynę konstytutywną chromosomów (głównie u roślin);
– prążki G – wybarwiane odczynnikiem Giemsy (metoda GTG) lub Wrighta (metoda GTW) po trawieniu proteolitycznym; uwidaczniają regiony bogate w adeninę i tyminę, dzięki czemu pozwalają nie tylko na identyfikację chromosomów, ale również na uzyskanie informacji o ogólnej organizacji DNA i genów;
– prążki R – uwidaczniane barwnikiem Giemsy po denaturacji termicznej; odwrotność prążków G; reprezentują charakterystyczne dla euchromatyny obszary bogate w cytozynę i guaninę;
– prążki N – występują głównie w obszarze organizatora jąderkowego NOR;
– prążki T– uzyskiwane w zastosowania barwnika Giemsy w podwyższonej temperaturze;
– Ag- NOR – to najczęściej stosowana metoda barwienia organizatorów jąderkowych; zastosowany azotan srebra wybarwia białka związane z aktywnym transkrypcyjnie DNA;
• Barwione fluorescencyjnie – z wykorzystaniem fluorochromów wykazujących powinowactwo do obszarów bogatych w pary AT lub GC. Uzyskany w ten sposób wzór można wzmocnić stosując dodatkowy barwnik (fluorochrom lub związek nie wykazujący fluorescencji):
– Prążki Q – najstarsza metoda, która wykorzystuje barwnik quinakrynę (atebrynę) dający seledynowe zabarwienie o różnej intensywności, widoczne w regionach bogatych w pary AT;
– CMA – barwienie DNA bogatego w pary GC antybiotykiem A3 oraz mitramycyną (żółto-czerwona fluorescencja); to odwrotność prążków DAPI;
– DAPI – DAPI- 4’-6-diamidyno-2-fenylindol selektywnie łączy się z sekwencjami bogatymi w pary AT dając jasnoniebieską barwę.
Stosowane w cytogenetyce nazewnictwo regulowane jest przez Międzynarodowy System Nazewnictwa Cytogenetycznego (ang. International System for Human Cytogenetic Nomenclature). Pozycję prążka na chromosomie określa się podając numer chromosomu, jego ramienia (ramię krótkie p lub ramię długie q), numer regionu oraz numer prążka. Obszary usytuowane w pobliżu centromeru oznacza się numerem 1. Kolejne liczby wyznaczają regiony oddalające się w kierunku telomerów.
Autor: Anna Kurcek
Literatura:
• Stanisława Rogalska, Jolanta Małuszyńska, Maria J. Olszewska „Posdtawy cytogenetyki roślin” Wydanie II uaktualnione; Wydawnictwo Naukowe PWN; Warszawa 2005;
• Wilhelm Grzesiak, Arkadiusz Marian Kawęcki „Genetyka Zwierząt – przewodnik do ćwiczeń”; Szczecin 1998;
• Maria Joanna Olszewska „Neocentromery. II. Molekularne czynniki niezbędne dla powstawania centromeru i neocentromeru”, Postępy Biologii Komórki Tom 35, nr. 2 (273 – 285); 2008;
• Ewa Pyłka-Gutowska „Biologia. Vademecum Maturzysty”; Wydawnictwo „Oświata”; Warszawa 2002;