
DNA – budowa chemiczna, wiązania, heliks, konformacje, badanie struktury, trójniciowe DNA.
Rys. 1. Budowa helisy DNA
DNA ma postać heliksu (inaczej „podwójnej helisy”). Zbudowane jest z deoksyrybonukleotydów. W skład pojedynczego deoksyrybonukletydu wchodzą: – zasada azotowa: adenina lub guanina (dwupierścieniowa puryna), cytozyna lub tymina (jednopierścieniowa pirymidyna) – 2-deoksyryboza (pentoza), – reszta fosforanowa.
Zasada azotowa połączona jest z pierścieniem pentozowym cukru wiązaniem N-glikozydowym. Natomiast deoksyrybonukletydy połączone są ze sobą wiązaniami fosfodiestrowymi. Wiązanie te występuje pomiędzy atomem 5’ węgla w reszcie cukrowej jednego deoksyrybonukleotydu, a atomem 3’ reszty cukrowej drugiego.
Deoksyryboza jest pochodną rybozy, w której grupa hydroksylowa połączona z węglem 2′ zastąpiona jest grupą wodorową. Grupa fosforanowa składa się z jednej, dwóch lub trzech reszt fosforanowych związanych z atomem 5′ reszty cukrowej. Nukleozydem nazywamy cząsteczkę, która składa sie jedynie z zasady azotowej i pentozy.
Pełne nazwy chemiczne czterech nukleotydów: – 2′-deoksyadenozyno-5′-trifosforan: dATP – 2′-deoksycytydyno-5′-trifosforan: dCTP – 2′-deoksyguanozyno-5′-trifosforan: dGTP – 2′-deoksytymidyno-5′-trifosforan: dTTP
Skąd się wziął heliks?
Zasady azotowe są nierozpuszczalne w wodzie o obojętnym pH, w przeciwieństwie do deoksyrybozy, która rozpuszcza się dobrze. W związku z tym cząsteczki DNA tworzą strukturę wewnątrz której znajdują się elementy nierozpuszczalne w wodzie, a na zewnątrz elementy rozpuszczalne (cukier i reszta fosforanowa). W warunkach fizjologicznych DNA ma postać regularnego heliksu. Podwójną helisę tworzą dwa polinukleotydy skierowane w przeciwnych kierunkach i razem zwinięte.
Odziaływania w obrębie heliksu: – wiązania wodorowe pomiędzy adenina i tyminą lub pomiędzy cytozyną i guaniną. Warto zauważyć, że w warunkach fizjologicznych tylko puryna i pirymidyna tworzą pary, – oddziaływania hydrofobowe między zasadami.
Opisana helisa to forma B, w komórce DNA występuje głównie w tej formie. Wiadomo jednak, że helisa jest na swój sposób elastyczna i może przyjmować różne kształty. Najważniejsze zmiany konformacji powoduje rotacja wokół wiązania β-N-glikozydowego oraz obrót wokół wiązania między atomami węgla 3′ i 4′ reszty cukrowej. Poza helisą typu B rozróżniamy również helisy: A, B’, C, C’, C”, D, E, T. Warto wiedzieć, że wszystkie te formy DNA są prawoskrętne. Znana jest jednak również forma lewoskrętna – jest to helisa Z.
Na powierzchnii helisy B rozróżniamy dwa rowki: większy i mniejszy. Również helisa A posiada rowki, mają one nieco inne wymiary niż w przypadku formy B. Forma Z charakteryzuje sie obecnością tylko jednego rowka.
Na dnie rowka białka wiążące DNA rozpoznają sekwencję nukleotydową DNA, ponieważ „wystają tam” grupy chemiczne zasad azotowych. Zmiany konformacji w obrębie DNA mogą zatem byc mechanizmem regulującym ekspresję genów.
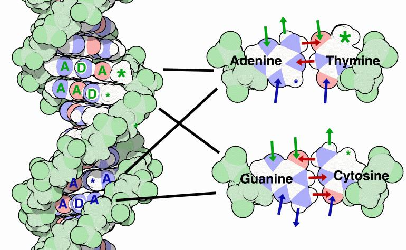 |
| Rys. 2. Struktura DNA, powstaje dzięki komplementarności par zasad |
Badanie struktury DNA za pomocą analizy wzoru przecinania
Badanie struktury DNA odbywa się w roztworze wodnym w warunkach fizjologicznych. Pierwszym etapem jest unieruchomienie cząsteczki DNA na płaskiej powierzchni. Zwykle odbywa się to na powierzchni kryształów fosforanu wapnia. Ponieważ jedna strona helisy jest „przyklejona” do powierzchni, dlatego też jest ona niedostępna dla czynników znajdujących się w roztworze. Druga zaś strona jest wyeksponowana. Im miejsce bardziej eksponowane tym bardziej narażone jest na przecinanie. Ze względu na budowę podwójnej helisy łatwo zauważyć, że miejsca takie powtarzają się co jeden pełny obrót, a jeśli tak to możemy w ten sposób określić długość skoku podwójnego heliksu. DNaza I jest enzymem najczęściej wykorzystywanym do przecinania DNA zaaobsorbowanego na stałych powierzchniach, podobnie jak kompleksy żelaza z EDTA. DNaza I charakteryzuje się stosunkowo niewielką specyficznością w stosunku do sekwencji zasad. Produkty powstałe w wyniku przecinania DNA denaturuje się (doprowadza do postaci jednoniciowej) i rozdziela za pomocą elektroforezy poliakrylamidowej. Skok podwójnego heliksu to odległości pomiędzy najintensywniejszymi prążkami.
Trójniciowe DNA
Pary typu Watsona-Cricka (A i T oraz G i C) nie są jedynym ułożeniem zasad spotykanym w obrębie DNA. Karst Hoogsteen wykazał, że w kryształach złożonych z adeniny i tyminy pary A-T wyglądają inaczej niż pary typu Watsona-Cricka. Pary Hoogsteena nie występują w naturalnym DNA czego główną przyczyną jest niestabilność pary G-C. Niemniej jednak wiedza na temat par typu Hoogsteena ma ogromne znaczenie dla opracowania leków oddziałujących w określonym miejscu genomu.
Literatura:
Biologia molekularna w medycynie. Elementy genetyki klinicznej. Praca zbiorowa pod redakcją J. Bala. Polskie Wydawnictwo Naukowe 2008.
Genetyka Molekularna. Praca zbiorowa pod redakcją Piotra Węgleńskiego. Polskie Wydawnictwo Naukowe 1995.
Genomy. Brown T.A. Polskie Wydawnictwo Naukowe 2001.