
Mimikra molekularna to zjawisko homologii i podobieństwa strukturalnego, serologicznego i funkcjonalnego pomiędzy antygenami drobnoustrojów a tkankami organizmu wyższego. Określa się ją jako zdolność indukowania przez struktury chemiczne patogenów, przeciwciał reagujących zarówno z antygenami gospodarza jak i antygenami drobnoustrojów chorobotwórczych, czyli tzw. przeciwciał krzyżowo-reagujących. U podstaw tego zjawiska leży podobieństwo pewnych struktur I-rzędowych antygenów gospodarza i bakterii. Podobieństwo to może wynikać z podobnej sekwencji aminokwasów lub podobnej konformacji cząsteczki i może występować u bardzo odrębnych grup organizmów. W takim wypadku odpowiedź immunologiczna przeciwko determinantom antygenowym występującym zarówno u gospodarza jak i patogenu może wywołać tkankowo-specyficzną odpowiedź immunologiczną, która może doprowadzić do reakcji autoimmunologicznej a w konsekwencji może wywołać zniszczenie komórek i tkanek, poprzez własne mechanizmy efektorowe gospodarza tj. układ komórek fagocytarnych czy układ komplementu.
Rola mimikry molekularnej w etiologii chorób
Mimikra molekularna uznawana jest za ważny czynnik decydujący o patogenności drobnoustrojów, wpływający na rozwój pewnych chorób autoimmunologicznych. Jest ona jedną ze strategii drobnoustrojów chorobotwórczych umożliwiającą wniknięcie patogenu do organizmu gospodarza, dotarcie do komórki docelowej, a następnie oddziaływanie na procesy wewnątrz- i zewnątrzkomórkowe w celu wymknięcia się spod kontroli układu immunologicznego gospodarza. Podobieństwo między antygenami drobnoustrojów takimi jak: węglowodany, lipopolisacharydy, glikolipidy i białka, a autoantygenami organizmu człowieka może doprowadzić do przekształcenia się łagodnej infekcji w chorobę autoimmunologiczną. Tolerancja immunologiczna na obce antygeny, lecz tożsame z własnymi strukturami powoduje, że bakteria nierozpoznana jako obcy czynnik może wnikać, migrować i namnażać się, prowadząc do zakażeń, a nawet wstrząsu septycznego. Przykładem jednostek chorobowych, których etiologia związana jest z zjawiskiem mimikry molekularnej są: gorączka reumatyczna, przewlekły gościec postępujący, pierwotna marskość żółciowa, choroba Gravesa- Basedowa i wiele innych.
Przykładowe jednostki chorobowe których etiologia związana jest z zjawiskiem mimikry molekularnej
Główną przyczyną gorączki reumatycznej są bakterie Gram dodatnie a dokładnie paciorkowce z grupy A, wywołujące również zakażenia górnych dróg oddechowych, mogące doprowadzić do licznych powikłań. Powikłania te związane są przede wszystkim z występowaniem podobieństw strukturalnych pomiędzy składnikami ściany komórkowej np. węglowodanami, mukopeptydami i białkami M, a ludzkimi tkankami. Mimikra cząsteczkowa jest również przyczyną przewlekłego gośćca postępującego. Antygenami bakteryjnymi które są związane z występowaniem tej jednostki chorobowej są białka szoku cieplnego. W organizmie człowieka również dochodzi do ekspresji białek szoku termicznego na powierzchni komórek w tkankach objętych procesem zapalnym. Mimikrę molekularną odkryto m.in. między białkiem Hsp60 z Escherichia coli a antygenami hsp człowieka, co wpływa na powstawanie autoprzeciwciał i stymulację limfocytów T. Co ciekawe – białka szoku cieplnego posiadają silne właściwości immunoregulacyjne oraz wykazują zdolności do wywołania odpowiedzi zapalnej, co przemawia za hipotezą, że białka Hsp mogą pośredniczyć miedzy infekcją a chorobą autoimmunologiczną. Głównym czynnikiem patogennym w pierwotnej marskości żółciowej jest pałeczka jelitowa Escherichia coli. Z przeprowadzonych badań wynika, że czynnikiem zapoczątkowującym rekcje autoimmunologiczną jest podobieństwo między podjednostką E2 dehydrogenazy pirogronianowej człowieka i Escherichia coli.
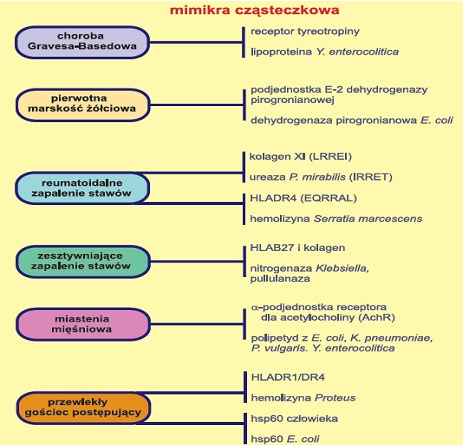
Rys.1 Rola mimikry cząsteczkowej w etiologii niektórych chorób (1).
Mimikra czynników translacyjnych
Proces translacji polega na odczytywaniu informacji o białku zapisanej w sekwencji matrycowego RNA z jednoczesnym tworzeniem łańcucha polipeptydowego. Proces ten dzieli się na trzy etapy: inicjację, elongację i terminację. Terminacja translacji związana jest z koniecznością rozpoznania kodonu stop i podjęcia przez rybosom i białka wspomagające stosownej reakcji skutkującej uwolnieniem z rybosomu powstającego łańcucha polipeptydowego. W terminacji translacji u Prokariota niezbędna jest obecność kodonu „stop” (UAG, UGA, UAA) oraz cytoplazmatycznych czynników uwalniających RF (ang. Release Factor). Czynniki RF to niewielkie białka, których funkcją jest oddziaływanie z miejscem A rybosomu (rozpoznają kodony „stop”) prowadzące do hydrolizy, tj. odłączenia łańcucha peptydowego od tRNA. Czynnik uwalniający zmienia specyficzność peptydylotransferazy – rybozymu bedącego elementem rybosomu, w ten sposób, że akceptorem aktywowanej reszty peptydylowej nie jest grupa aminowa ale cząsteczka wody. Uwolniony łańcuch polipeptydowy opuszcza rybosom, uwolnione zostają tRNA, mRNA i czynniki RF. Ostatecznie rybosom dysocjuje na podjednostki 30S i 50S i może rozpocząć się synteza innej cząsteczki białkowej. Czynniki RF1 (rozpoznaje kodony UAA i UAG) i RF2 (rozpoznaje kodony UGA i UAA) wykazują zjawisko molekularnej mimikry przypominając strukturalnie tRNA. Dzięki takiemu podobieństwu możliwe jest ich przyłączenie do kodonu stop w miejscu A na rybosomie.
Autor: Beata Rola
Literatura:
1. Witkowska D., Bartyś A., Gamian A. 2009. Białka osłony komórkowej pałeczek jelitowych i ich udział w patogenności oraz odporności przeciwbakteryjnej. Postepy Hig Med Dosw. 63: 176-199.
2. Witkowska D. 1999. Mimikra cząsteczkowa jako czynnik patogenności bakterii. Postepy Hig Med Dosw. 53: 554-558.
3. Stanisławska J., Interewicz B., Olszewski W., 2003. Odpowiedź leukocytów gospodarza na antygeny bakteryjne. Post. Mikrobiol. 42, 3, 301-317.
4. Bugla-Płoskońska G. 2008. Organizacja struktur zewnętrznych bakterii gram-ujemnych a proces aktywacji układu immunologicznego. Post. Mikrobiol. 47, 3, 191-197.
5. Guilherme L., Köhler K.F., Postol E. 2011. Genes, autoimmunity and pathogenesis of rheumatic heart disease. Ann Pediatr Cardiol. 4: 13–21.